from the Aegean islands Crete and Karpathos
(Amphibia, Salientia, Ranidae)
PETER BEERLI, HANSJÜRG HOTZ
Zoologisches Museum, Universität Zürich-Irchel, Winterthurerstrasse 190,
CH-8057 Zürich, Switzerland
HEINZ G. TUNNER, SUSANNA HEPPICH
Biozentrum, Universität Wien, Althanstrasse 14,
A-1090 Wien, Austria
THOMAS UZZELL
Department of Ecology, Ethology and Evolution, University of Illinois,
515 Morrill Hall, 505 South Goodwin Avenue, Urbana, Illinois 61801, USA
Recent analyses using protein electrophoresis suggest that the water frog populations of the Rana esculenta group occurring on two of the Aegean islands, Crete and Karpathos, are both genetically differentiated from the other western Palearctic water frogs at the species level (Beerli 1993). We here describe these two new species.
Distribution. Known only from Crete, where it is widely distributed; our samples are all from below 100 m above sea level.
 Fig. 1. Rana cretensis: adult female (paratype, MHNG 2543.91) of 72 mm
body length from Petros river, Crete.
Fig. 1. Rana cretensis: adult female (paratype, MHNG 2543.91) of 72 mm
body length from Petros river, Crete.
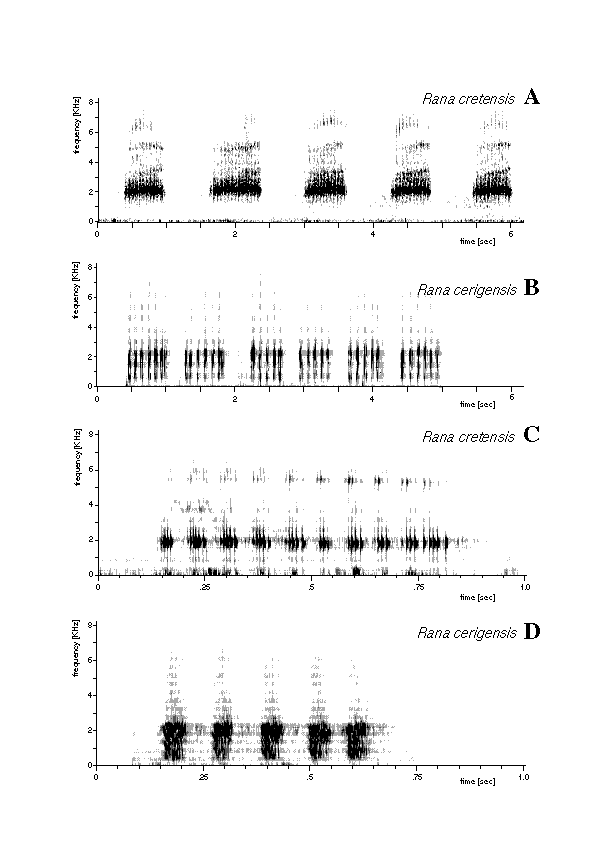 Fig. 2. Sonograms of the advertisement call of an individual Rana
cretensis from Petros river, Crete, water temperature 20ëC, air
temperature 26ëC (A,C), and of an individual Rana cerigensis from
Olympos, Karpathos, water temperature 20°C, air temperature 18°C
(B,D).
Fig. 2. Sonograms of the advertisement call of an individual Rana
cretensis from Petros river, Crete, water temperature 20ëC, air
temperature 26ëC (A,C), and of an individual Rana cerigensis from
Olympos, Karpathos, water temperature 20°C, air temperature 18°C
(B,D).
Distribution. Karpathos, possibly Rhodos.
 Fig. 3. Rana cerigensis: adult female (paratype, MHNG 2543.93) of 76 mm
body length from Olympos, Karpathos.
Fig. 3. Rana cerigensis: adult female (paratype, MHNG 2543.93) of 76 mm
body length from Olympos, Karpathos.
Several islands in the Aegean Sea have been isolated for a long time, up to five million years. Some of these are inhabited by water frogs of the western Palearctic Rana esculenta group. For these frogs, as for other amphibians, the salt water surrounding the islands is an effective dispersal barrier, because the skin of these fresh-water animals is readily permeable to both salt and water. Barring introduction by humans, the geologically determined isolation time of such islands is, therefore, the minimal isolation time between the respective frog populations and thus their minimal genetic divergence time.
The island Crete, although isolated during most of the Miocene, became connected to the surrounding mainlands when the Mediterranean basin dried up 6 million years ago (salinity crisis); it subsequently became completely isolated again 5 million years ago (at the end of the Messinian period) by filling of the dry Mediterranean basin following the re-opening of the Strait of Gibraltar (Hsü et al. 1977). Analysis of the complex tectonic movements in the Aegean region and fossil data suggest that the island Karpathos, between Crete and Rhodos, was connected to Rhodos and the Anatolian mainland in the early Pliocene, and became completely isolated in middle Pliocene (Meulenkamp 1985).
This scenario indicates that the water frogs living on Crete diverged from the other western Palearctic water frogs a minimum of 5 million years and a maximum of 6 million years ago, and that the water frogs of Karpathos diverged from Anatolian relatives about 3 million years ago. Genetic data obtained by protein electrophoresis are consistent with these datings (Beerli 1993).
The population on Crete has similar genetic distances to the three geographically nearest mainland water frog taxa, Rana ridibunda and Rana epeirotica from the Balkan peninsula, and Rana bedriagae 3[4] from Anatolia (Table 1). These distances are among the lowest found between Rana cretensis and any other western Palearctic water frog taxon (Table 1); but they are similar to or higher than genetic distances between European taxon pairs that are known by sympatry not to be conspecific (Rana ridibunda-Rana lessonae, Rana ridibunda-Rana shqiperica, Rana ridibunda-Rana epeirotica; Beerli 1993, Hotz, Beerli and Uzzell, unpublished data). This and the large number of fixed allelic differences (minimum in pairwise comparisons: 11 of 31 loci; 7 of 31 loci fixed for unique alleles) clearly indicate that Rana cretensis is differentiated from the other western Palearctic water frogs at the species level.
Table 1. Genetic distances of Rana cretensis and Rana cerigensis from the presently known western Palearctic water frog taxa (protein electrophoresis, 31 loci; data from Beerli 1993). Values are means of Nei's (1972) standard genetic distance as modified by Hillis (1984), and of Cavalli-Sforza and Edwards' (1967) chord distance.
Rana cretensis Rana cerigensisBy genetic distance measures (Table 1), Rana cretensis is closest to the western Greek Rana epeirotica. Nevertheless, the differences in genetic distances between Rana cretensis and several other species, and in numbers of fixed allelic differences distinguishing Rana cretensis from these species (including the geographically nearest mainland taxa, Rana ridibunda in Greece and Rana bedriagae in Anatolia) are too small to determine its closest relative reliably. A very conservative cladistically-derived tree of relationships as well as a robust maximum likelihood tree both place Rana cretensis, Rana epeirotica, Rana shqiperica, Rana lessonae, and the Rana ridibunda stock (Rana cerigensis, Rana bedriagae, Rana ridibunda) together in an unresolved group (Beerli 1993). The conspicuous centromeric fluorescence shown by Rana cretensis chromosomes following Actinomycin D/33258 Hoechst double staining resembles that shown by double-stained chromosomes of the Rana ridibunda stock (cf. Heppich et al. 1982; Bucci et al. 1990; Tunner and Heppich-Tunner 1991); whether this reflects convergence or common inheritance remains to be tested. In our phylogenetic analyses, Rana perezi and Rana saharica consistently form an outgroup pair.Nei / Cavalli-Sforza Nei / Cavalli-Sforza
Hillis and Edwards Hillis and Edwards
Rana cretensis - - 0.54 0.58
Rhodos population 0.55 0.59 0.12 0.29
Rana bedriagae Anatolia 0.58 0.58 0.32 0.44
Rana ridibunda Balkan 0.65 0.62 0.27 0.44
Rana ridibunda central Europe 0.70 0.64 0.30 0.46
Rana epeirotica 0.53 0.57 0.49 0.56
Rana shqiperica 0.66 0.63 0.49 0.56
Rana lessonae central Europe 0.89 0.69 0.51 0.57
Rana perezi 0.80 0.67 0.73 0.65
Rana saharica Morocco 1.35 0.77 0.95 0.71
Compared to Rana cretensis, Rana cerigensis is less markedly different from Rana bedriagae, the genetically and geographically closest mainland taxon (Table 1; fixed allelic differences at 2 of 31 loci). Speciation within the Rana ridibunda stock needs further study through both molecular genetics and crossing experiments. In all our cladistic and phenetic analyses of protein electrophoretic data, however, Rana cerigensis, while clearly belonging to the Rana ridibunda stock, is the most divergent member of it, and is clustered with the animals from the neighboring island Rhodos as a distinctive unit. If Rana bedriagae is recognized as a species distinct from Rana ridibunda (cf. Schneider et al. 1992), which is consistent with although not required by our present genetic data (Beerli 1993; unpublished results), then Rana cerigensis, which is genetically more distinct, should also be recognized.
Although our analyses of the electrophoretic data cluster the frogs from Karpathos and Rhodos, which suggests including the Rhodos population within Rana cerigensis, the geological history clusters Rhodos with the Anatolian mainland: it became separated from Karpathos ~3 million years ago and from the Anatolian mainland in early Pleistocene, ~1.8 million years ago (Meulenkamp 1985). If these datings are correct, the Karpathos and Rhodos populations do not share an independent evolutionary history. The phylogenetic clustering results from alleles present in the Rhodos population (at 30 of 31 loci) that are shared (exclusively so for sAAT l, AHH c, and EST-5 e) with the Karpathos population; at five of these loci (sAAT, AHH, CA-2, mIDH, MPI), however, the Rhodos population segregates for alternative alleles that occur in Rana bedriagae on the adjacent Anatolian mainland. If the geological history is correct, these data may reflect alleles of ancestral populations that subsequently became extinct in Anatolia. Additional molecular data are needed to determine the evolutionary history and taxonomic status of the water frogs on Rhodos.
There are few water frog populations on the dry island of Karpathos, and population sizes appear to be modest. Given deforestation, scarcity of surface water, and degradation of aquatic habitat by human activity, the long-term survival of Rana cerigensis on Karpathos is uncertain: there is serious danger of its becoming extinct.
Our view of the phyletic radiation of the western Palearctic water frog group has changed substantially in the past two decades. Two species were recognized by Mertens and Wermuth (1960), but evolutionary studies evoked by the hemiclonal reproduction that characterizes widespread natural hybrid lineages (reviewed by Graf and Polls Pelaz 1989, Gönther 1990) have resulted in recognition of at least 9 species today. Most of our current ideas on phylogenetic relationships among western Palearctic water frogs have been shaped by genetic data using biochemical and molecular markers. The two new species described here, like the discovery of Rana shqiperica and of Rana epeirotica (Hotz and Uzzell 1982, Tunner and Heppich 1982), and the recognition of Rana perezi and of Rana saharica as distinct species (Graf et al. 1977, Uzzell 1982; unpublished data), reflect the importance of molecular data in alpha taxonomy.
BUCCI, S., M. RAGGHIANTI, G. MANCINO, L. BERGER, H. HOTZ AND T. UZZELL. 1990. Lampbrush and mitotic chromosomes of the hemiclonally reproducing hybrid Rana esculenta and its parental species. Journal of Experimental Zoology 255(1): 37-56.
CAVALLI-SFORZA, L. L. AND A. W. F. EDWARDS. 1967. Phylogenetic analysis: models and estimation procedures. Evolution 21(3): 550-570.
GRAF, J.-D., F. KARCH AND M.-C. MOREILLON. 1977. Biochemical variation in the Rana esculenta complex: A new hybrid form related to Rana perezi and Rana ridibunda. Experientia 33(12): 1582-1584.
GRAF, J.-D. AND M. POLLS PELAZ. 1989. Evolutionary genetics of the Rana esculenta complex. In: Dawley, R.M. and J.P. Bogart (Ed.): Evolution and ecology of unisexual vertebrates. New York State Museum Bulletin 466: 289-302.
GÜNTHER, R. 1990. Die Wasserfrösche Europas (Anura - Froschlurche). Die Neue Brehm-Bücherei (Wittenberg Lutherstadt) 600: 288 pp.
HEPPICH, S., H. G. TUNNER AND J. GREILHUBER. 1982. Premeiotic chromosome doubling after genome elimination during spermatogenesis of the species hybrid Rana esculenta. Theoretical and applied Genetics 61: 101-104.
HILLIS, D. M. 1984. Misuse and modification of Nei's genetic distance. Systematic Zoology 33(2): 238-240.
HOTZ, H. 1983. Genic diversity among water frog genomes inherited with and without recombination. Ph.D. Dissertation, UniversitÙt Zörich: 136 pp.
HOTZ, H. AND T. UZZELL. 1982. Biochemically detected sympatry of two water frog species: two different cases in the Adriatic Balkans (Amphibia, Ranidae). Proceedings of the Academy of Natural Sciences of Philadelphia 134: 50-79.
HSÜ, K. J., L. MONTADERT, D. BERNOULLI, M. B. CITA, A. ERICKSON, R. E. GARRISON, R. B. KIDD, F. MÈLIERÉS, C. MÜLLER AND R. WRIGHT. 1977. History of Mediterranean salinity crisis. Nature 267: 399-403.
MERTENS, R. AND H. WERMUTH. 1960. Die Amphibien und Reptilien Europas (Dritte Liste, nach dem Stand vom 1. Januar 1960). Senckenberg-Buch 38. W. Kramer, Frankfurt am Main: XII+264 pp.
MEULENKAMP, J. E. 1985. Aspects of the late Cenozoic evolution of the Aegean region. In: Stanley, D. J. and F. C. Wezel (Ed.): Geological evolution of the Mediterranean basin. Springer, New York: 307-321.
NEI, M. 1972. Genetic distance between populations. American Naturalist 106(949): 283-292.
NISHIOKA, M. AND M. SUMIDA. 1992. Biochemical differentiation of pond frogs distributed in the Palearctic region. Scientific Report of the Laboratory for Amphibian Biology, Hiroshima University, 11(2): 71-108.
SCHNEIDER, H., U. SINSCH AND E. NEVO. 1992. The lake frogs in Israel represent a new species. Zoologischer Anzeiger 228(1/2): 97-106.
TUNNER, H. G. AND S. HEPPICH. 1982. A genetic analysis of water frogs from Greece: evidence for the existence of a cryptic species. Zeitschrift för zoologische Systematik und Evolutionsforschung 20(3): 209-223.
TUNNER, H. G. AND S. HEPPICH-TUNNER. 1991. Genome exclusion and two strategies of chromosome duplication in oogenesis of a hybrid frog. Naturwissenschaften 78(1): 32-34.
UZZELL, T. 1982. Immunological relationship of western Palearctic water frogs (Salientia: Ranidae). Amphibia-Reptilia 3(2/3): 135-143.